

0 引言
【研究意义】我国小麦栽培面积和总产量均居谷物第一位,但禾谷丝核菌引起的小麦纹枯病则导致小麦不同程度减产。特别是随着秸秆粉碎入田耕作制度的实行,小麦纹枯病的发病率逐年提高[1]。截至目前尚无对小麦纹枯病菌具有有效抗性的小麦品种[2]。与不同的防治策略相比,利用环境友好的微生物进行生物防治被认为是防治小麦纹枯病的一种有效途径[3]。【前人研究进展】作为最大的放线菌属,链霉菌占土壤放线菌总数的50%[4]。这些链霉菌产生丰富多样的次级代谢产物,不仅可以促进植物生长,还具有对多种植物病原菌的生物防治潜力[5
1 材料与方法
1.1 材料
试验在新疆维吾尔自治区农业科学院新疆作物化学调控工程技术研究中心温室进行。3个小麦品种矮抗58、百农307和周麦22由新疆农业科学院新疆作物化学调控工程技术研究中心提供。
葡萄糖大豆蛋白胨酵母提取物培养基(GPY):葡萄糖20 g,大豆蛋白胨5 g,酵母提取物5 g,去离子水1 000 mL。
马铃薯葡萄糖琼脂培养基(PDA):马铃薯200 g,葡萄糖20 g,琼脂粉15 g,去离子水1 000 mL。
1.2 方法
1.2.1 链霉菌HU2014发酵液的制备
保存的链霉菌HU2014在PDA固体培养基上活化后,用打孔器将菌株打成直径5 mm的菌饼,将其接种到GPY液体培养基里,于25℃、150 r/min下摇床培养12 d。将得到的发酵原液先用筛绢初滤,再用0.25 mm针头滤芯过滤得无细胞发酵液,稀释500倍后对小麦进行灌根处理。
1.2.2 试验设计
将3个供试小麦品种催芽后播种在花盆中(每盆15株,12 cm×12 cm×9 cm),于温室中培养(25℃ / 20℃)±1℃,光照/黑暗:14 h /10 h,相对湿度65%左右,正常浇灌。待长出2叶1心的小麦苗后,按照每盆100 mL进行灌根处理。灌根前为0 h,灌根后2、6、12、24、48和96 h,分别采集100 mg叶片放入1.5 mL离心管中,立刻投入液氮中冷冻3~5 min,再放置于-80℃超低温冰箱中保存。
1.2.3 RNA提取,cDNA合成及实时荧光定量PCR
采用Trizol法提取小麦叶片总RNA。参照Prime ScriptTM RT reagent kit with gDNA Eraser(TaKaRa,中国,大连)中说明书的方法合成第一链cDNA。以1 mg总RNA为模板,RT Primer Mix为引物,在200 mL的RNase-Free离心管中进行反应。以Actin作为内参基因,通过Applied biosystems 7500 Real-Time PCR System,检测各个设定基因的转录量,比较小麦经供试药剂诱导后的基因表达量差异,反应程序:95℃ 30 s; 95℃ 5 s; 60℃ 34s,40个循环。试验重复3次,结果以小麦目标基因与内参基因(Actin)的拷贝数比值表示。采用2-ΔΔCt方法进行相对表达量的计算分析,采用SPSS18软件(IBM,美国,纽约)进行显著性分析。定量PCR引物设计参照Genbank数据库。表1
表1 定量PCR引物序列
Tab.1
| 引物 Gene name | 上下游引物 Forward and reverse primers (5'-3') |
|---|---|
| TaActin | GGACCTCACGGATAATCTAATG TGACCATCAGGCATCTCA |
| TaPR1 | AACCTCGGCGTCTTCATCA TTTACTCGCTCGGTCCCTCT |
| TaPR2TGCCGTTGCTCTCTTCAT ATGCCCTTGGACCTGTAGA | |
| TaPR3 | AGAGATAAGCAAGGCCACGTC GGTTGCTCACCAGGTCCTTC |
| TaPR5 | ACAGCTACGCCAAGGACGAC CGCGTCCTAATCTAAGGGCAG |
| PAL | CCAATGTTCTGTCCGTCCT GCTGCTTCAATCTGTCCAG |
| LOX | AGGCAACTACATCTACGCTTC GCCATCAACACCAGAGTCA |
2 结果与分析
2.1 HU2014诱导AK58抗性基因表达
研究表明,在HU2014发酵液处理小麦品种AK58后,TaPR1基因表达量在随后的几个接种时间与对照相比,显著降低。TaPR2基因在6 h与对照相比显著升高,其余时间点均显著降低。TaPR3基因表达量与对照相比,除2 h与对照相比无显著差异外,其余时间点均表达显著降低。TaPR5基因的表达量呈现降低趋势(除72 h无显著差异外)。LOX基因在6 h与对照相比显著升高,其余时间点均显著降低。PAL基因的表达在48和96 h时与对照比无显著差异,其余时间点均表达显著降低。图1
图1
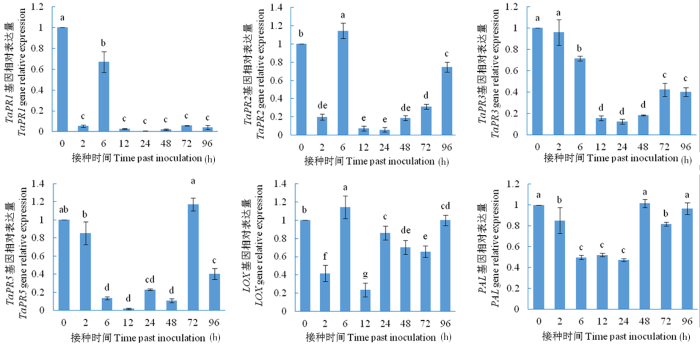
图1
链霉菌HU2014对小麦AK58的抗性基因表达
Fig.1
The strain HU2014 on the expression of resistance genes in wheat (cultivar AK58) plant
2.2 HU2014诱导BN307抗性基因表达
研究表明,在HU2014发酵液处理小麦品种BN307后,6个基因均在不同时间点显著表达。TaPR1基因在24 h后的积累最高,与对照相比,倍数值为6.95。TaPR2基因的表达在24 h 时与对照相比达到最高水平,其倍数值为55.70,其余时间点均表达显著降低。TaPR3基因表达量在随后的几个时间点与对照相比均显著降低。TaPR5基因的表达量在24 h出现小高峰,此后急剧降低,但到96 h积累最高,其倍数值为3.33。LOX基因在6和24 h均表达量最高,其余时间点均显著降低。PAL基因的表达在24和48 h时积累最高,其余时间点均表达显著降低。图2
图2
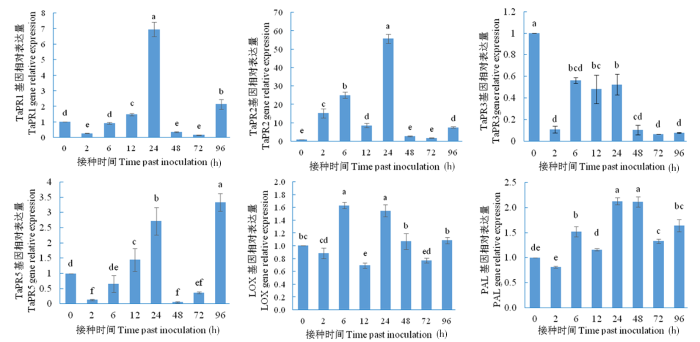
图2
链霉菌HU2014对小麦BN307的抗性基因表达
Fig.2
The strain HU2014 on the expression of resistance genes in wheat (cultivar BN307) plant
2.3 HU2014诱导ZM22抗性基因表达
研究表明,在HU2014发酵液处理小麦品种ZM22后,除TaPR1基因表达量在对照点时间后均显著降低外,其余5个基因均在不同的处理时间点显著升高。TaPR2基因的表达在12 h 时与对照相比达到最高水平,其倍数值为105.47,其余时间点均表达显著降低。TaPR3基因表达量在2 h最高,其余时间点均显著降低。TaPR5基因的表达量在6 h出现小高峰,此后急剧降低,但到12 h积累最高,其倍数值为98.67。LOX基因在6 h表达量最高,其余时间点均显著降低。PAL基因的表达在24和48 h积累较高,其余时间点均表达显著降低。图3
图3
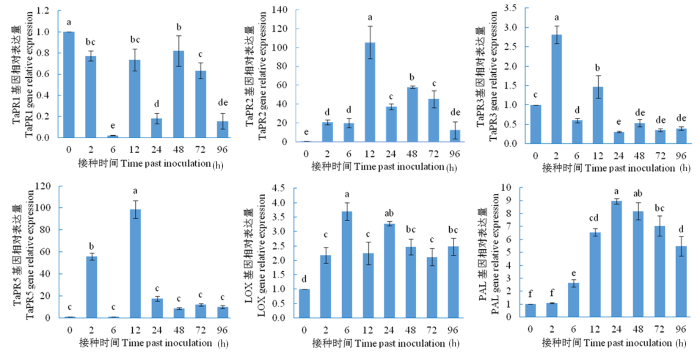
图3
链霉菌HU2014对小麦ZM22的抗性基因表达
Fig.3
The strain HU2014 on the expression of resistance genes in wheat (cultivar ZM22) plant
3 讨论
3.1 链霉菌除了通过产生拮抗化合物直接抑制病原菌外,还可以通过激活宿主植物的抗性途径间接为其植物宿主提供保护[22-
3.2 苯丙氨酸解氨酶(PAL)是苯丙酸合成途径的限速酶,脂氧合酶(LOX)是茉莉酸途径的关键酶。水杨酸途径基因(PAL)和茉莉酸途径基因(LOX)在试验研究中也做了相应的表达分析。HU2014发酵液处理在不同程度上诱导了PAL和LOX酶的转录谱。该菌可能通过诱导小麦的水杨酸和(或)茉莉酸途径基因的表达,提高小麦的抗病性。相关研究也证明了这点,例如链霉菌N2中提取出来的抗真菌霉素N2处理水稻幼苗,PAL的表达量与对照组相比显著提高[34];在病原菌和不吸水链霉菌公主岭变种(农抗“769”)的双重诱导下,大豆中PAL的表达量比对照组提高了95.22%[35];经链霉菌JD211菌种的发酵液处理后的水稻叶片中LOX活性较对照组提高了40.85%[36]。
另外,从研究中可知,经HU2014无细胞发酵液处理后,TaPR1基因在AK58和ZM22中下调,TaPR3基因在AK58和BN307中下调。ZM22中TaPR2、TaPR3、TaPR5、PAL和LOX的最高表达水平高于AK58和BN307,而BN307中TaPR1的最高表达水平高于AK58和ZM22。以上结果表明同一抗病基因在不同小麦品种的表达谱中表现有差异,可能与小麦遗传背景有关。
试验基于HU2014的发酵液通过灌根的盆栽方式进行苗期小麦的抗性基因表达分析,而小麦在不同的生育阶段,基因表达存在差异,小麦的抗病性表现会有不同。因此,对小麦整个生育期的抗病性能还有待进一步研究。其次,该菌的发酵液对小麦根际土壤的影响也需要进行相关的研究,以揭示其诱导抗病性的相关机制。
4 结论
链霉菌HU2014诱导小麦抗性基因TaPR1、TaPR2、TaPR3、TaPR5、PAL和LOX的表达。TaPR1基因在AK58和ZM22中下调,TaPR3基因在AK58和BN307中下调。ZM22中TaPR2、TaPR3、TaPR5、PAL和LOX的最高表达水平高于AK58和BN307,而BN307中TaPR1的最高表达水平高于AK58和ZM22。链霉菌HU2014可以诱导小麦系统性获得抗性和诱导性系统抗性,并且这种诱导抗性作用可能与水杨酸和茉莉酸合成途径有关。
参考文献
小麦、玉米两熟秸秆还田土壤中6种有机酸对小麦纹枯病的化感作用
[J].
Allelopathy of six organic acids on wheat sheath blight in the soil of winter wheat-summer maize double cropping straw returning system
[J].
小麦纹枯病的发生及防治技术
[J].
The occurrence and control technology of wheat sheath blight
[J].
小麦纹枯病化学和生物防治研究进展
[J].
Advances in chemical and biological control of wheat sharp eyespot
[J].
List of prokaryotic names with standing in nomenclature (LPSN) moves to the DSMZ
[J].
The taxonomy of Streptomyces and related Genera
[J].
Evaluation of broad-spectrum streptomyces sp. for plant growth promotion traits in chickpea (Cicer arietinum L.)
[J].
Streptomyces-induced resistance against oak powdery mildew involves host plant responses in defense, photosynthesis, and secondary metabolism pathways
[J].
Plant growth-promoting abilities and biocontrol efficacy of Streptomyces sp. UPMRS4 against Pyricularia oryzae
[J].
Defense priming of tomato plants by Streptomyces metabolites to combat Corynespora cassiicola and Pseudomonas syringae infestations
[J].
Methyl salicylate is a critical mobile signal for plant systemic acquired resistance
[J].
Evaluation of the local isolate Streptomyces kanamyceticus strain Tikrit-5 in control of gray rot disease on eggplant caused by Botrytis cinerea
[J].
褪黑素诱导小豆抗锈病机理的初步研究
[J].
A preliminary study on the mechanisms of melatonin-induced rust resistance of adzuki bean
[J].
Which plant proteins are involved in antiviral defense?review on in vivo and in vitro activities of selected plant proteins against viruses
[J].
Ecology, Epidemiology, and Control of Plant Viruses
[M].
柑桔褐斑病菌侵染对不同抗性柑桔种质防御酶活性的影响
[J].
Effects of Alternariaalternata infection on defense enzyme activity of different resistant citrus germplasm
[J].
链霉菌JD211发酵液对水稻防御稻瘟病菌诱导抗性的作用
[J].
Effect of Streptomyces JD211 fermentation products on the induced resistance to Magnaporthe grisea in rice
[J].
Transcriptomic analysis reveals the mechanism of MtLOX24 in response to methyl jasmonate stress in Medicago truncatula
[J].
Lipidomic and transcriptomic profiles of glycerophospholipid metabolism during Hemerocallis citrina Baroni flowering
[J].
Soybean (Glycine max L.) lipoxygenase 1 (LOX 1) is modulated by nitric oxide and hydrogen sulfide: an in vitro approach
[J].
实时荧光定量PCR技术研究进展及其应用
[J].
Research progress and application of real-time fluorescence quantitative PCR
[J].
Endophytic Actinobacteria induce defense pathways in Arabidopsis thaliana
[J].
Plant behavior upon contact with streptomycetes
[J].
Plant-growth-promoting rhizobacteria
[J].
The sterol-binding activity of PATHOGENESIS-RELATED PROTEIN 1 reveals the mode of action of an antimicrobial protein
[J].
Subcellular localization of Arabidopsis pathogenesis-related 1 (PR1) protein
[J].
Characterization of the biogenic volatile organic compounds (BVOCs) and analysis of the PR1 molecular marker in Vitis vinifera L. inoculated with the nematode Xiphinema index
[J].
Co-ordinated regulation of chitinase and β-1, 3-glucanase in bean leaves
[J].
Structures of an active-site mutant of a plant 1, 3-β-glucanase in complex with oligosaccharide products of hydrolysis
[J].
Pathogenesis related defence functions of plant chitinases and β-1, 3-glucanases
[J].
Antiphytopathogenic and plant growth promoting attributes of Bacillus strains isolated from rhizospheric soil of chickpea
[J].
Drought-tolerant Streptomyces pactum Act12 assist phytoremediation of cadmium-contaminated soil by Amaranthus hypochondriacus: great potential application in arid/semi-arid areas
[J].
Endoplasmic reticulum-related E3 ubiquitin ligases: key regulators of plant growth and stress responses
[J].
Induced defense responses against Rhizoctonia solani in rice seedling by a novel antifungalmycin N2 from Streptomyces sp. N2
[J].
不吸水链霉菌公主岭变种诱导大豆抗病性与根际土壤微环境的变化
[J].
Changes of soybean disease resistance and the rhizosphere micro-environment induced by Streptomyces gongzhulingensis n.var
[J].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}