

0 引言
【研究意义】种子异型性(seed heteromorphism)或称种子异态是指同一植株能够产生不同形状或者不同萌发行为的种子的现象[1]。具有种子异型现象的植物多为一年生草本植物并且多分布于荒漠地和盐碱地等环境变化迅速的地区[2]。种子异型作为植物对于多变环境的一种适应性特征[3],常在形状、颜色、大小和结构等方面表现出不同,在生态行为方面异型性又通常与种子的扩散、繁殖、萌发和休眠等特性联系在一起[4⇓-6],荒漠环境复杂多变,干旱少雨,且在不同时间内温度、湿度以及降水的变化显著。种子异型性便是荒漠植物为适应异质环境而进化出的一种特殊机制,而种子不同的扩散方式和发芽机制与荒漠一年生植物能否顺利存活息息相关。【前人研究结果】藜科是具种子异型性植物的第二大科,藜属(Chenopodium)、盐角草属(Salicornia)、猪毛菜属(Salsola)、碱蓬属(Suaeda)、滨藜属(Atriplex)等[7]。藜科植物一般情况下能产生2种或3种在颜色、大小、着生方式等方面多有不同的异型种子,具备两种散布结构,一种散布结构是种子上包裹的苞片,另一种则是种子宿存的花被片,部分花被片带有翅或刺等附属物,且存在休眠和非休眠两种类型的种子[8]。【本研究切入点】蔷薇猪毛菜为藜科猪毛菜属的一年生草本植物,主要分布在哈萨克斯坦、西伯利亚等中亚国家及地区中,我国蔷薇猪毛菜主要分布在新疆阿勒泰、乌鲁木齐、阜康、奇台等地;多生长于海拔700~1 100 m的平原盐碱地、山前洪积扇砾石荒漠及低山山坡;具有较强的抗干旱、耐盐碱能力,对于维持荒漠生态平衡和维持天然牧草资源方面都有着重要的作用[9]。目前,有关猪毛菜属植物的研究已有报道,但对蔷薇猪毛菜的种子异型性与适应机制还尚不清楚。【拟解决的关键问题】以准噶尔荒漠蔷薇猪毛菜为材料,研究猪毛菜植株个体大小、繁殖分配,种子输出与萌发特性,为分析种子异型植物提供数据支持。
1 材料与方法
1.1 材料
研究区位于准噶尔盆地南缘(43°48’44"N,87°40’26"E),主要建群种为小蓬(Nanophytonerinaceum)、蔷薇猪毛菜、紫翅猪毛菜(Salsola affinis)、散枝猪毛菜(Salsola bracchita)等,盖度为20%~30%。该地区属于中温带荒漠区,春季秋季湿润,夏季炎热干旱,冬季时间长且气温低;年蒸发量大于降水量,年平均日较差、地面温度日较差变化很大。材料蔷薇猪毛菜为藜科一年生草本植物,果实为胞果,内含有1粒种子,胚螺旋状。
1.2 方法
1.2.1 样本的采集与植株个体大小的确定
2019年10月在蔷薇猪毛菜果实成熟期采集植株样本,根据其高度、冠幅、分枝数等大致分成大、中、小3个等级,总共采集86株。在取样前,对植物形态、颜色和种子的颜色、扩散等观察记录。取样时,用小十字镐小心挖取完整植株,每一株样本单独放置于密封袋内保存,确保植株结构的完整性。但野外环境存在干扰因素或其他误差,最终以自然晾干后的植株干重作为蔷薇猪毛菜个体大小的划分标准。
1.2.2 种子类型的划分
随机抽取蔷薇猪毛菜20株,根据种子宿存花被和翅的形态、种子形态、颜色、着生方式等对种子分类。判断种子类型后选取不同类型种子各50粒,用游标卡尺测量其直径;种子重量的测定以每种类型种子100粒为1组,用万分之一天平称重,重复4次。
1.2.3 结实
选取不同等级的蔷薇猪毛菜植株各20株,统计株高、节数、分枝数、不同类型种子数目,计算不同类型种子的输出比例;称量种子重量和植株干重,计算繁殖分配。
1.2.4 不同变温条件下种子萌发
将不同类型种子的萌发试验设置为每组25粒,4个重复。将种子放置于垫有2层滤纸的培养皿中并加入10 mL蒸馏水。将种子分别放入5℃/15℃、10℃/20℃、15℃/25℃、20℃/30℃、25℃/35℃(黑暗/光照为12 h/12 h)的变温培养箱中进行萌发试验,第1 d,每2 h观察1次,第2 d开始,每天观察1次,2 d加1次蒸馏水,试验连续观察14 d,每天记录种子萌发状态。判断种子萌发的标准是,突破种皮且种子打开螺旋。
1.3 数据处理
使用Excel软件对初始数据进行整合处理,使用SPSS20.0单因素方差分析(P=0.05)对植株生物量、种子生物量、繁殖分配、萌发率等进行差异显著性分析,使用Origin18.0对相关数据进行绘图。
2 结果与分析
2.1 蔷薇猪毛菜植株形态特征
研究表明,干重分为3个等级,3个等级干重具有极显著的差异(P<0.05)。Ⅰ级个体最轻,单个植株平均重量为(1.110±0.100) g;Ⅲ级个体最重,单个植株平均重量为(8.941±1.680) g,随着植株干重的增加,植株的株高、节数、分枝数明显增多,都具有极显著差异(P<0.05)。I级植株个体矮小,分枝数少;Ⅲ级为大型植株,个体高大且分枝数较多。表1
表1 蔷薇猪毛菜不同大小个体的形态特征(平均值±标准误)
Table 1
| 植株等级 Class of plant | 植株干重 Plant biomass (g) | 株高 Plant height (cm) | 节数 No. of nodes | 分枝数 No. of branches |
|---|---|---|---|---|
| Ⅰ | 1.110±0.100c | 28.617±1.430c | 24.460±1.499c | 2.960±0.643c |
| Ⅱ | 3.708±0.235b | 43.556±1.005b | 45.400±1.993b | 9.630±0.856b |
| Ⅲ | 8.941±1.680a | 49.533±3.979a | 55.000±2.031a | 18.500±2.377a |
注:同一列内不同的小写字母标记的值之间差异显著(P<0.05),下同
Note: Within a column different superscript letters mean difference at P < 0.05,the same as below
2.2 蔷薇猪毛菜种子形态
图1
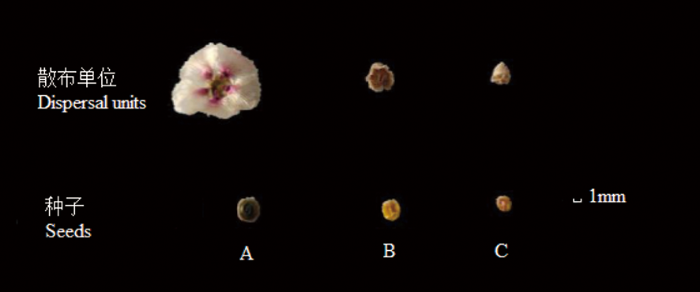
图1
蔷薇猪毛菜的3种类型种子形态特征
Fig.1
Morphological characteristics of three types of seeds of Salsola rosacea
表2 蔷薇猪毛菜3种类型种子特征比较(平均值±标准误)
Table 2
| 种子类型 Seed types | 颜色 Color | 形状 Shape | 翅 Wings | 长 Length (mm) | 种子单粒重 Seed single weight (mg) | 着生方式 Orientation of seed |
|---|---|---|---|---|---|---|
| A | 绿色 | 圆形 | 长 | 2.451±0.036a | 3.124±0.057a | 横生 |
| B | 黄色 | 圆形 | 短 | 1.700±0.022b | 1.725±0.032b | 横生 |
| C | 黄色 | 圆形 | 无 | 1.348±0.063c | 1.522±0.026c | 横生 |
2.3 蔷薇猪毛菜依赖个体大小的繁殖分配
研究表明,个体大小与繁殖体生物量:植株干重、株高和三种类型繁殖体生物量在不同等级别间都差异显著(P<0.05),且在同一种群中随着植株个体的增大而增大,表现出对个体大小的依赖。
Ⅰ级植株总繁殖分配比例在12.256左右,Ⅱ级植株总繁殖分配比例在13.132左右,Ⅲ级植株总繁殖分配比例在14.432左右,随着个体的增大总繁殖分配而增大;在试验统计中发现,最小的植株重量只有0.28 g时,植株仍有繁殖体输出,总繁殖分配达到8.507%。
3种植株大小中A型繁殖体和B、C两种类型繁殖体在繁殖分配比例上差异显著(P<0.05)。Ⅰ级植株和Ⅲ级中A型繁殖体繁殖分配比例最高,B型繁殖体所占比例最低;而在Ⅱ级植株中C型繁殖体所占繁殖分配比例最低。不同植株大小的A、B、C型繁殖体的分配比例上存在显著差异;A型随植株个体的增大而增大且在三种繁殖体类型中所占繁殖分配比例均为最高,在10.214%~11.561%内;B型繁殖分配比例亦随个体的增大而增大,变化范围为0.876%~1.397%;C型繁殖体的繁殖分配比例随个体大小变化不明显,不依赖个体大小而变化。
相同等级的植株在3种繁殖体的分配比例上显现差异性,在Ⅰ级和Ⅲ级植株中A型繁殖体的繁殖分配比例最大,B型繁殖体所获投资最少;对Ⅱ级植株来说,C型所占繁殖分配比例最小。表3
表3 蔷薇猪毛菜不同大小个体的繁殖分配(平均值±标准误)
Table 3
| 植株等级 Class | 繁殖体总生物量 Total (g) | 繁殖分配 Reproductive allocation (%) | 总繁殖分配 Total (%) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| A | B | C | A | B | C | ||||
| Ⅰ | 0.113±0.010Ac | 0.012±0.003Bc | 0.013±0.002Bc | 10.214±0.422Aa | 0.876±0.190Ba | 1.166±0.167Ba | 12.256±0.453b | ||
| Ⅱ | 0.397±0.034Ab | 0.042±0.007Bb | 0.035±0.006Bb | 11.069±0.343Aa | 1.121±0.163Ba | 0.943±0.148Ba | 13.132±0.283ab | ||
| Ⅲ | 0.968±0.099Aa | 0.118±0.019Ba | 0.121±0.009Ba | 11.561±0.340Aa | 1.397±0.166Ba | 1.474±0.096Ba | 14.432±0.252a | ||
注:同一列内不同的小写字母表示相同类型种子不同植株等级之间差异显著(P<0.05),同一行内不同的大写字母表示相同植株等级不同种子类型之间差异显著(P<0.05),下同
Note: Different lowercase letters in the same column indicated significant differences between different plant classes of the same type of seed (P< 0.05). Different uppercase letters in the same row indicated significant differences between different plant classes of the same type of seed (P< 0.05),the same as below
2.4 蔷薇猪毛菜依赖个体大小的种子输出
研究表明,不同等级的植株个体中A、B、C 3种类型种子的数目和输出比例存在差异。
其中种子总数(P<0.05)、A型种子数(P<0.05)、B型种子数(P<0.05)、C型种子数(P<0.05)在3个不同级别的植株中均具有极显著的差异(P<0.05),且随植株等级的增加而增加。
3种类型种子输出比例的变化有所不同,其中A型种子输出比例随植株等级的增加逐渐减低,B型种的输出比例随植株个体的增大而增大,表现出对个体大小的依赖;C型种输出比例基本保持稳定,无显著变化。
同一级别植株的种子不同类型的种子输出也存在差异。对于I级植株,3种类型种子输出存在显著差异(P<0.05),从高到低依次为A型种子、C型种子、B型种子;对于Ⅱ级Ⅲ级植株,A型种子输出最多,B型和C型种无显著差异。表4
表4 蔷薇猪毛菜不同大小个体的种子输出变化
Table 4
| 植株等级 Class of plant | 种子总数 Total | 种子数量 No. of seeds | 种子输出 Seed output (%) | ||||
|---|---|---|---|---|---|---|---|
| A | B | C | A | B | C | ||
| Ⅰ | 49.739± 4.155c | 38.609± 3.401Ac | 3.913± 0.829Bc | 7.217± 1.274Bc | 77.931± 2.570Aa | 7.457± 1.634Cb | 14.612± 1.906Ba |
| Ⅱ | 186.539± 16.266b | 135.692± 11.439Ab | 26.692± 3.830Bb | 24.154± 3.641Bb | 73.532± 2.571Aab | 13.887± 1.354Ba | 12.581± 1.587Ba |
| Ⅲ | 451.167± 53.639a | 301.500± 36.846Aa | 71.000± 9.957Ba | 78.667± 8.515Ba | 66.762± 1.019Ab | 15.564± 1.049Ba | 17.673± 0.930Ba |
2.5 蔷薇猪毛菜异型种子萌发特性
图2
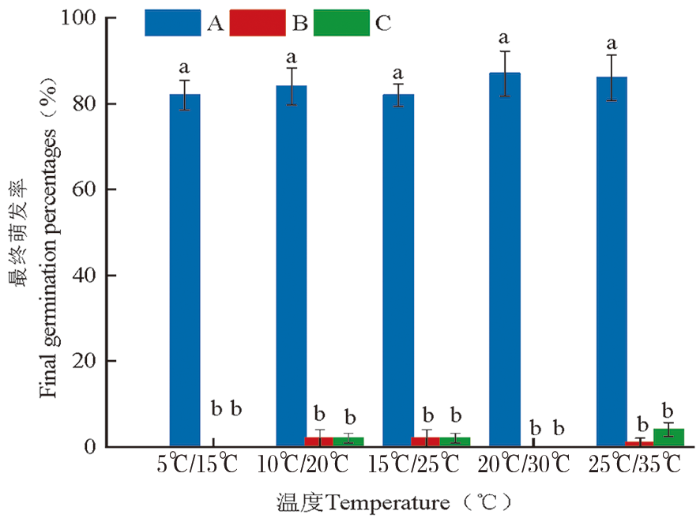
图2
蔷薇猪毛菜3种类型种子在不同变温条件下的最终萌发率变化
Fig.2
The final germination percentages (%±SE) of three types of seeds of Salsola rosacea at different alternating temperatures
图3
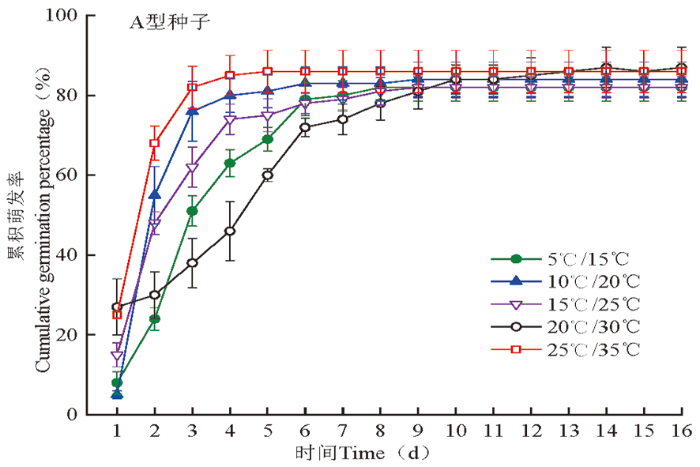
图3
蔷薇猪毛菜A型种子在不同变温条件下的累积萌发率变化
Fig.3
The cumulative germination percentages (%±SE) of A type of seeds of Salsola rosacea at different alternating temperatures
种子类型对蔷薇猪毛菜种子的萌发具有极显著影响(P<0.01),而温度对其没有显著影响(P>0.05),交互作用也影响不显著(P>0.05)。表5
表5 蔷薇猪毛菜不同类型种子与不同温度范围对萌发影响的双因素方差
Table 5
| 偏差来源 Source | 自由度 df | 均方 Meansquare | F | P |
|---|---|---|---|---|
| 种子类型 Type of seed | 2 | 45 817.867 | 1 641.564 | 0 |
| 温度 Temperature | 4 | 14.267 | 0.511 | 0.728 |
| 种子类型×温度 Type of seed×Temperature | 8 | 10.867 | 0.389 | 0.921 |
3 讨论
植物的个体大小的表型可塑性是植物适应异质环境的重要途径[13],可能导致植物在自然种群中出现差异[14]。蔷薇猪毛菜按个体大小来分,将其分为3种不同等级的植株:Ⅰ级、Ⅱ级、Ⅲ级。在蔷薇猪毛菜野外采样观察中发现,植株的高度,生物量都存在显著性差异。大型植株的生物量是小型植株生物量的8倍,高度和节数是小型植株的2倍,分枝数达到小型植株的9倍。与已报道的异型性植物紫翅猪毛菜[15]与散枝猪毛菜[16]相似。可能是因为植物在个体大小方面的表型可塑性能够使其更容易在多变的环境下生存定居。一般情况下造成种群个体呈现不同大小现象和大小依赖下的繁殖分配的主要因素是种内与种间的竞争,资源限制和遗传差异;而决定这种依赖性的关键是植株本身对于资源获取和繁殖分配的生理权衡[17]。
植物的繁殖特性被认为是植物表型可塑性的重要特征[18]。合理地利用和分配有限的资源以及权衡各种功能一直是植物繁殖策略研究的重要内容[19]。植物为了适应多变环境会产生形态大小不同的种子以便其在遇到适宜的条件时适合度能够呈现出“表现最大化”[20-21]。在胁迫的自然环境下生存的植物的繁殖分配比例也许会随着环境波动而有所下降,也有可能不发生改变[22]。不同大小种子的比例及其数量会受到植株个体大小的影响[23],也存在于其他藜科植物中[15-16]。紫翅猪毛菜的繁殖分配比例会因为植株个体的增大而使A型种子的占比递增,B型种子占比逐步降低,但C型种子比例不产生显著差异[15],随着植株大小的变化,有翅的种子占比基本都随之变化,而不具翅的种子基本输出稳定。研究发现蔷薇猪毛菜随植株等级的增加,A型和B型种子所占比例也因此而产生变化,而C型种子输出比例稳定,这与前人的结果相似。
不同大小形态的种子比例的变化会改变其后代的传播能力及萌发特性,也可能使种群的适合度受到影响[24-25]。异型种子在散布能力、形态和萌发特性等方面均存在差异[26]。异型种子中一种种子形态会具有较高的扩散能力,通常很少或没有休眠,而另一种种子形态具有较低(或没有)扩散能力,具有较高的休眠[27-28]。研究蔷薇猪毛菜产生的3种种子中,A型种子具长翅可以利用风作为媒介散布到较远的地方逃离原始栖息地严酷的环境,更远的散布单位的比例在种群扩张中也具有重要意义[29]。A型种不仅可萌发温度范围广而且温度和降水合适就能迅速萌发;B、C型种子则具短翅或不具翅只能在离母体不远处传播或保留在母株之上,在一定程度上避免了与A型种竞争。同时,B和C型种表现出一定的休眠特性,存在延迟发芽现象。就延迟发芽而言,前一年生产的一部分种子不能完全发芽,会保留在土壤种子库中[30-31]。种子库在确保植物种群的持久性方面发挥着重要作用[32-33],可作为避免种群灭亡的储备力量,有助于物种未来的遗传变异。种子异型性和结实以及传播模式的某种组合可能是植物在自然生境中的独特适应策略。不同类型种子的特性决定了植物具备与之相对应的生物学功能,而不同的萌发机制和扩散策略又共同形成了一种混合繁殖策略[34]。
4 结论
4.1
蔷薇猪毛菜有3种类型种子,具有不同的散布方式。其中A型种子主要分布于植株的上部,长翅使其散布范围更广;B型种子大部分生长于植株的中下部,短翅使其较容易与母株脱离;C型种子主要分布于植株的中下部或分布在果枝的最顶端及基部,生理上可能存在休眠迹象,在土壤中形成种子库。在经历生理后熟之后有可能具备萌发的能力。
4.2
绿色(A型)种子萌发速度快,不存在休眠,而黄色种子(B型和C型)萌发率低且存在休眠现象。绿色(A型)种子采取“机会型”的萌发策略,适应温度范围广,条件适宜能迅速萌发,而黄色种子采取“保护型”的萌发策略,具有休眠特性。
参考文献
The evolutionary ecology of seed heteromorphism
[J].
种子异型性及其生态意义的研究进展
[J].
种子异型性是指同一植株产生不同形状或行为种子的现象。根据异型种子在植株上的生长位置, 种子异型性可划分为地上下结实性和地上种子异型性两类。此现象已在26科129属292种被子植物中报道。异型性种子植物主要分布于干旱半干旱区、荒漠和盐渍土地区等干扰强烈的环境, 在菊科和藜科中最为常见, 主要出现在一年生植物中。种子异型性在避免密集负效应、减弱同胞子代间的竞争、采取两头下注策略以适应时空异质性环境等方面具有重要的进化生态意义。该文系统总结了国内外种子异型性的研究工作, 主要内容包括: 1)种子异型性的概念、类型和种类, 2)具有异型种子植物的生境和生活型, 3)异型种子的生态学特性, 4)种子异型性的理论模型, 5)种子异型性的生态意义。在综述文献的基础上, 对今后的研究进行了展望。针对国内外的研究现状, 提出两点建议: 1)系统调查具有种子异型性现象的植物种类, 摸清其生物学特性; 2)确定研究种子异型性现象的模式植物, 从生态学、生理学和分子生物学等学科角度来研究种子异型性的个体发育机制及分子调控机理。
Review of research on seed heteromorphism and its ecological significance
[J].<FONT face=Verdana>Seed heteromorphism is the production of seeds of different form or behavior by single individuals. According to the relative position of seeds on the plant, seed heteromorphism can be divided into amphicarpy and heterodiaspory. Seed heteromorphism has been reported in 26 families, 129 genera and 292 species of angiosperms. Most heteromorphic plants studied are annuals, often species in Asteraceae and Chenopodiaceae faced with stochastic environments such as arid, semiarid, desert and saline soil. Seed heteromorphism is generally considered to play an important role in escaping from the negative effect of crowding, reducing sib competition and adapting to environment by following bet-hedging strategy. We review and analyze recent advances in seed heteromorphism research, with emphases on 1) conception and types of seed heteromorphism and the species of the heteromorphic plants, 2) habitats and life types of the heteromorphic plants, 3) ecological characteristics of heteromorphic seeds, 4) theoretical model of seed heteromorphism and 5) ecological significance of seed heteromorphism. In addition, we discuss prospects for further research in this area and suggest 1) species with seed heteromorphism should be investigated and their biological characteristics should be studied and 2) the model plant of seed heteromorphism needs to be selected and ecological, physiological and molecular biological methods should be used to reveal the ontogenetic and molecular controlling mechanisms.</FONT>
How does seed heteromorphism influence the life history stages of Atriplex sagittata (Chenopodiaceae)
[J].
植物种子二形性(多形性)研究进展
[J].
Advances in plant seed dimorphism (or polymorphism) research
[J].
Seed polymorphism, dormancy and germination of Salsola affinis (Chenopodiaceae), a dominant desert annual inhabiting Junggar Basin of Xinjiang, China
[J].
Germination strategies of halophyte seeds under salinity
[J].
果翅、盐分及干旱胁迫对白梭梭种子萌发的影响
[J].
Effect of winged perianth, salt & drought stress on germination of Halaxylonpersicum
[J].
Ecological consequences and ontogeny of seed heteromorphism
[J].
The shapes and sizes of seeds
[J].
Population ecology of halophyte seeds
[J].
表型可塑性与外来植物的入侵能力
[J].外来植物的入侵能力与其性状之间的关系是入侵生态学中的基本问题之一。成功的入侵种常常能占据多样化的生境,并以广幅的环境耐受性为特征。遗传分化(包括生态型分化)和表型可塑性是广布性物种适应变化、异质性生境的两种不同但并不矛盾和排斥的策略。越来越多的实验证据表明,表型可塑性具有确定的遗传基础,本身是一种可以独立进化的性状。许多入侵种遗传多样性比较低,但同时又占据了广阔的地理分布区和多样化的生境,表型可塑性可能在这些物种的入侵成功和随后的扩散中起到了关键作用。本文首先介绍表型可塑性的含义,简述表型可塑性和生物适应的关系,然后从理论分析和实验证据两个方面论述了表型可塑性与外来植物入侵能力的相关性,最后针对进一步的研究工作进行了讨论。当然,并非所有入侵种的成功都能归因于表型可塑性,作者认为对于那些遗传多样性比较低同时又占据多样化生境的入侵种,表型可塑性和入侵能力的正相关可能是一条普遍法则,而非特例。
Phenotypic plasticity and invasiveness of alien plants
[J].How the traits of invasive plants determine their invasiveness is one of the major issues in invasion biology. Invasive plants usually have broad ecological amplitudes, and hence can exploit a great diversity of habitats. Genetic differentiation and phenotypic plasticity are two major strategies that invasive plants can use to invade broad geographical areas and diverse ecosystems. Phenotypic plasticity is a trait that has a certain genetic basis and can evolve independently. Although some of the invasive plants have low genetic variation, they can still invade diverse habitats. Phenotypic plasticity may play a critical role in invasion of these species into new environments. In this review, the concept of phenotypic plasticity and its significance for adaptation were introduced. The relationship between phenotypic plasticity and the invasiveness of alien plants was discussed, supported by theoretical analysis and published experimental evidence.We also suggest some issues that might be addressed in future studies for better understanding of the mechanisms of successful invasion with reference to phenotypic plasticity. Although successful invasion cannot be explained by phenotypic plasticity alone, the positive correlation between invasiveness and phenotypic plasticity seems to be a rule rather than an exception for those invasive species with low genetic diversity but a broad geographic distribution.
紫翅猪毛菜的种子多型性及其结实格局
[J].紫翅猪毛菜(Salsola affinis)主要分布于新疆准噶尔荒漠, 具有很强的抗干旱和耐盐碱能力, 其种子具有多型现象。我们对紫翅猪毛菜的种子形态、萌发特性及结实格局进行了观测研究。结果表明: (1) 紫翅猪毛菜3种类型的种子在形状、大小、颜色、着生方式及包被其花被片背部是否具翅方面均有明显不同; (2) 3种类型的种子具有不同的散布特性和萌发行为: A型种子容易从母体脱落, 发达的果翅能够借助风进行远距离传播; B、C型种子成熟时紧连母体, 不易脱落。在变温下A型种子和B型种子能够快速萌发, 且B型种子比A型种子的萌发速度快; 而C型种子萌发缓慢, 最终萌发率小于10%, 表明其处于休眠状态; (3)随植株个体的增大, A型种子所占比例逐渐增多, 由0.43%增加到51.07%; B型种子所占比例逐渐减少, 由65.80%减少到18.06%; C型种子所占比例没有显著变化, 约为30–35%。紫翅猪毛菜的种子多型性以及种子输出的灵活性有利于其在荒漠异质环境中成功定居。对种子多型性及其结实格局的研究将为探讨其生态适应机制及生活史进化研究提供科学依据。
Seed polymorphism and fruit-set patterns of Salsola affinis
[J].<em>Salsola affinis</em>, a dominant desert annual inhabiting the Junggar Basin of Xinjiang, produces three types of seeds which differ in morphology and ecological properties. To better understand how <em>S. affinis</em> is adapted to its desert habitat, we studied its seed morphs, germination characteristics and fruit-set patterns. Having lignified perianths with long wings, Type A seeds can easily be dispersed to a long distance by wind, whereas type B and type C seeds were tightly attached to the mother plants because of short wings or absence of wings. Seeds were germinated in incubators at three alternating temperature regimes. Both Type A and Type B seeds could germinate rapidly at three alternating temperature regimes, although Type B seeds germinated more quickly. Type C seeds germinated slowly with germination rate less than 10%, indicating that they were dormant. With the increase of the size of the individual parent plant, the ratio of Type A increased from 0.43% to 51.07%, while Type B decreased from 65.8% to 18.06%, but Type C kept steady (30–35%). The success of <em>Salsola affinis</em> in the desert might be partly attributed to its heterocarpy and associated plastic response to heterogeneous environmental conditions.
散枝猪毛菜的果实多型性及个体大小依赖的繁殖输出
[J].
Studyon fruit polymorphismand size-dependent reproductive output of Salsola bracchita
[J].
Size dependent reproduction in Australian alpine ranunculus
[J].
Phenotypic plasticity of vegetative and reproductive traits in monoecious and dioecious populations of Sagittaria latifolia (Alismataceae): a clonal aquatic plant
[J].
Variations in seed size within populations of Silene dioica (L.) Clairv in relation to habitat
[J].
Phenotypic plasticity for fitness components in species of contrasting ecological breadth
[J].
Phenotypic plasticity in Polygonumpersicaria II. Norms of reaction to soil moisture ecological breadth and the maintenance of genetic diversity
[J].Adaptive phenotypic plasticity is the predicted evolutionary response to fine-grained fluctuation in major environmental factors, such as soil moisture in plant habitats. This study examines genotypes from two natural populations of Polygonum persicaria, one from a relatively homogeneous, moderately moist site, and one from a site in which severe drought and root flooding occur within single growth seasons. Norms of reaction (phenotypic response curves) were determined for a random sample of eight and ten cloned genotypes, respectively, from each of the populations over a controlled moisture gradient ranging from drought to flooding.© 1993 The Society for the Study of Evolution.
Effect of Seed dimorphism on the density-dependent dynamics of experimental populations of Atriplex triangularis (Chenopodiaceae)
[J].
Selection of offspring size at independence and other size-versus-number strategies
[J].
The evolutionary ecology of seed heteromorphism
[J].
Comparison of germination and seed bank dynamics of dimorphic seeds of the cold desert halophyte Suaedacorniculata subsp. Mongolica
[J].
Phenotypic plasticity and bet-hedging in a heterocarpic winter annual/spring ephemeral cold desert species of Brassicaceae
[J].
Diaspore dispersal ability and degree of dormancy in heteromorphic species of cold deserts of northwest China: a review
[J].
Is farther seed dispersal better Spatial patterns of offspring mortality in three rainforest tree species with different dispersal abilities
[J].
Bet-hedging germination in annual plants: a sound empirical test of the theoretical foundations
[J].
Within-and among-year germination in Sonoran Desert winter annuals: bet hedging and predictive germination in a variable environment
[J].In variable environments, organisms must have strategies to ensure fitness as conditions change. For plants, germination can time emergence with favourable conditions for later growth and reproduction (predictive germination), spread the risk of unfavourable conditions (bet hedging) or both (integrated strategies). Here we explored the adaptive value of within- and among-year germination timing for 12 species of Sonoran Desert winter annual plants. We parameterised models with long-term demographic data to predict optimal germination fractions and compared them to observed germination. At both temporal scales we found that bet hedging is beneficial and that predicted optimal strategies corresponded well with observed germination. We also found substantial fitness benefits to varying germination timing, suggesting some degree of predictive germination in nature. However, predictive germination was imperfect, calling for some degree of bet hedging. Together, our results suggest that desert winter annuals have integrated strategies combining both predictive plasticity and bet hedging.© 2016 John Wiley & Sons Ltd/CNRS.
Studies on the feasibility of re-creating chalk grassland vegetation on ex-arable land. I. The potential roles of the seed bank and the seed rain
[J].
Seed germination cues and the importance of the soil seed bank across an environmental gradient in the Serengeti
[J].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}